Вто рич ная по лость тела. Полости тела животных
Тип Кольчатые черви
Общая характеристика типа Кольчатые черви (кольчецы)
Общая характеристика типа
Кольчатые черви (кольчецы) - крупный тип (около 9 тыс. видов) высших свободножи вущих морских, пресноводных и почвенных животных, имеющих более сложную организацию, чем плоские и круглые черви. Это в первую очередь относятся к морским многощетинковым червям, которые являются узловой группой в эволюции высших беспозвоночных: от их древних предков произошли моллюски и членистоногие.
Главнейшие прогрессивные черты строения кольчецов следующие:
1. Тело состоит из многочисленных (5-800) сегментов (колец). Сегментация выражается не только во внешней, но и во внутренней организации, в повторяемости многих внутренних органов, что повышает выживаемость животного при частичном повреждении тела.
2. Группы сходных по строению и функциям сегментов у мно-гощетинковых червей объединены в отделы тела - головной, туловищный и анальную лопасть. Головной отдел образовался путем слияния нескольких передних сегментов. У малощетинковых червей сегментация тела однородная.
3. Полость тела вторичная, или целом, выстланный целомичес-ким эпителием. В каждом сегменте целом представлен двумя изолированными мешками, заполненными целомической жидкостью.

Рис 11.7. Головной конец нереиды: I1-глаза; 2 - щупальца; 3 -усики; 4 - параподии с пучками щетинок.
4. Кожно-мускульный мешок состоит из тонкой эластичной кутикулы, расположенных под ней однослойного эпителия и двух мышечных слоев: наружного -кольцевого, и внутреннего - сильно развитого продольного.
5. Впервые появившиеся специализированные органы движения - параподии - представляют собой боковые двулопаст-ные выросты стенок тела туловищных сегментов, в которые заходит целом. Обе лопасти (спинная и брюшная) несут большее или меньшее количество щетинок (рис. 11.7). У малощетинковых червей параподии отсутствуют, имеются только пучки с немногочисленными щетинками.
6. В пищеварительной системе, имеющей три отдела, передняя кишка сильно дифференцирована на ряд органов (рот, глотку, пищевод, зоб, желудок).
7. Впервые развившаяся кровеносная система замкнутая. Она состоит из крупных продольных спинного и брюшного сосудов, соединенных в каждом сегменте кольцевыми сосудами (рис. 11.8). Движение крови осуществляется за счет перекачивающей деятельности сократимых участков спинного, реже кольцевых сосудов. В плазме крови содержатся дыхательные пигменты, близкие к гемоглобину, благодаря которым кольчецы заселили местообитания с самым различным содержанием кислорода.

8. Органы дыхания у многощетинковых червей -жабры; это тонкостенные листовидные, перистые или кустистые наружные выросты части спинных лопастей параподии, пронизанные кровеносными сосудами. Малощетинковые черви дышат всей поверхностью тела.
9. Органы выделения - попарно расположенные в каждом сегменте метанефридии, выводящие конечные продукты жизнедеятельности из полостной жидкости. Воронка метанефри-дия находится в целоме одного сегмента, а идущий от нее короткий каналец открывается наружу в последующем сегменте (см. рис. 11.8,6).
10. Нервная система ганглионарного типа. Она состоит из парных надглоточных и подглоточных ганглиев, соединенных нервными стволами в окологлоточное нервное кольцо, и многих пар ганглиев брюшной нервной цепочки, по одной паре в каждом сегменте (рис. 11.8, а). Органы чувств разнообразны: зрения (у многощетинковых червей), осязания, химического чувства, равновесия.
11. Подавляющее большинство кольчецов- раздельнополые животные, реже гермафродиты. Половые железы развиваются либо под целомическим эпителием во всех туловищных сегментах (у многощетинковых червей), либо только в некоторых (у малощетинковых червей). У многощетинковых червей половые клетки через разрывы целомического эпителия поступают в жидкость целома, откуда выводятся в воду специальными половыми воронками или метанефридиями. У большинства водных кольчецов оплодотворение внешнее, у почвенных форм - внутреннее. Развитие с метаморфозом (у многощетинковых червей) либо прямое (у малощетинковых червей, пиявок). Некоторые виды кольчецов, кроме полового, размножаются и бесполым способом (фрагментацией тела с последующей регенерацией недостающих частей). Тип Кольчатые черви подразделяется на три класса - Многощетинковые, Малощетинковые и Пиявки.
Тип Кольчатые черви. Общая характеристика
Главными характерными признаками кольчатых червей являются:
-вторичная, или целомическая, полость тела;
Появление кровеносной и дыхательной систем;
Выделительная система в виде метанефридиев.
Краткая характеристика
|
Среда обитания |
Морские и пресноводные, наземные и подземные животные |
|
|
Строение тела |
Тело вытянутое, червеобразное, метамерного строения. Двусторонняя симметрия. Трехслойные. У многощетинковых имеются параподии |
|
|
Покровы тела |
Кутикула. В каждом членике 8 или более щетинок для передвижения. В коже много желез. В кожно-мускульном мешке продольные и поперечные мышцы | |
|
Полость тела |
Вторичная полость тела - целом, заполнена жидкостью, выполняющей роль гидроскелета | |
|
Пищеварительная система |
Рот, глотка, пищевод, зоб, желудок, кишечник, анальное отверстие | |
|
Органы дыхания |
Дыхание всей поверхностью тела. У многощетинковых имеются наружные жабры | |
|
Кровеносная система |
Замкнутая. Один круг кровообращения. Сердца нет. Кровь красная | |
|
Пара трубочек в каждом метамере - метанефридии | ||
|
Нервная система |
Окологлоточное нервное кольцо, брюшная нервная цепочка лестничного типа | |
|
Органы чувств |
Осязательные и светочувствительные клетки, у многощетинковых имеются глаза | |
|
Половая система и развитие |
Гермафродиты. Оплодотворение перекрестное. Развитие без метаморфоза. Оплодотворение внутреннее. Многощетинковые раздельнополые, оплодотворение наружное, развитие с метаморфозом | |
Класс малощетинковые

Класс Малощетинковые черви объединяет 4-5 тыс. видов. Длина их тела колеблется от 0,5 мм до 3 м.
Аудиофрагмент "Класс Малощетинковые черви" (00:54)
Внутреннее строение дождевого червя
Покровы тела и мышцы. Кожа червя состоит из одного слоя покровных клеток. Среди них есть клетки, выделяющие слизь. Под кожей расположены кольцевые и продольные мышцы. При сокращении кольцевых мышц тело дождевого червя удлиняется, утончается и продвигается вперед. При сокращении продольных мышц задняя часть подтягивается к передней. Передвижение происходит волнообразно.
Виртуальная лабораторная работа

Полость тела. Как мы уже знаем, дождевой червь относится к трёхслойным животным. Его тело состоит по существу из двух трубок, вложенных одна в другую. Наружная трубка представляет собой стенку тела, а внутренняя – стенку пищеварительного тракта. Полость тела, выстланная изнутри слоем клеток , расположена между ними. В полостной жидкости (она придаёт телу упругость) находятся внутренние органы.
Пищеварительная
система.
Пищеварительный
тракт начинается ртом, затем следуют
глотка, пищевод, зоб, желудок, кишечник
и анальное отверстие.
Кровеносная система. Кровеносная система предназначена для перемещения внутри тела кислорода, углекислого газа, питательных и других веществ. У дождевого червя кровь не выливается свободно в полость тела, а движется только внутри сосудов. Такая кровеносная система называетсязамкнутой . Кровеносная система состоит из двух главных сосудов : спинного и брюшного. По спинному кровь течёт вперед, по брюшному – назад. В области пищевода эти сосуды соединены кольцевыми сосудами, называемыми «сердцами». Они имеют мышечные стенки, с помощью которых нагнетают кровь в брюшной сосуд. Ко всем органам и к стенкам тела отходят мелкие кровеносные сосуды.
Органы дыхания.
Органы дыхания. Органы дыхания у дождевого червя отсутствуют. Дыхание осуществляется через влажную кожу, пронизанную кровеносными сосудами.
Выделительная система. Выделительная система представлена парными органами (выделительными трубочками), расположенными в каждом сегменте тела. С помощью выделительной системы организм удаляет излишки воды и других веществ.
Нервная система. Нервная система состоит из окологлоточного нервного кольца и брюшной нервной цепочки с утолщениями в каждом сегменте, от которых отходят нервы. Окологлоточное кольцо состоит из надглоточного и подглоточного нервных узлов, соединённых кольцевой перемычкой. Специальных органов чувств нет, но чувствительные клетки в коже позволяют дождевому червю ощущать прикосновение и отличать свет от тьмы. Возникшее в этих клетках возбуждение передаётся по нервным волокнам в ближайший нервный узел, а оттуда по другим нервным волокнам – к мышцам, что вызывает их сокращение. Таким образом нервная система осуществляет ответ организма на раздражение (рефлекс).
2. Размножение и развитие
Дождевой червь способен размножаться как бесполым, так и половым способами. При бесполом размножении тело дождевого червя распадается на две части, а затем, путём регенерации, каждая из них «достраивает» недостающие отделы тела.
3. Значение дождевых червей
Кольчатые черви служат важными звеньями в цепях питания природных биогеоценозов. Дождевыми червями, например, питаются кроты, ежи, лягушки, жабы, птицы. Другими червями (например, трубочником ) – рыбы (карп, карась).
Дождевые черви приносят огромную пользу человеку, разрыхляя почву. Они делают её более пористой, более доступной для проникновения воздуха и воды, чем облегчается рост растений и увеличивается урожай сельскохозяйственных культур. Роясь в земле, черви проглатывают кусочки почвы, измельчают их и выбрасывают вместе с органическим веществом.
Чарлз Дарвин заинтересовался дождевыми червями ещё в молодые годы. В 1837 г. он сделал в Лондонском геологическом обществе доклад на тему: «Об образовании почвенного слоя», в котором изложил теорию, согласно которой частицы почвы всё время выносятся дождевыми червями из глубины на поверхность, благодаря чему предметы, лежащие на земле, оказываются по прошествии немногих лет на глубине 6–10 см под дёрном. Таким образом, весь почвенный слой оказывается прошедшим через желудок дождевых червей.
Чарлза
Дарвина поразили следующие наблюдения:
черви тянут в норку листья, ухватив их
за вершину (не за черешок), поэтому лист
оказывает наименьшее сопротивление
при затаскивании в норку(ведь вершина
листа уже его основания). А вот сосновые
иглы черви всегда тянут за черешок, то
есть общее для сдвоенных игл основание.
И в этом случае предмет оказывает
наименьшее сопротивление. Дарвин в
своих опытах подкладывал червям
вырезанные из бумаги треугольники, и
те втягивали их наиболее целесообразным
способом: за один из острых углов.
Дальнейшие исследования показали, что
ведущую роль в этом процессе играют
рефлексы.
Дождевые черви способны к обучению. Их помещали в Т-образный лабиринт: в наиболее длинный коридор, образующий основание буквы Т. Когда черви доползали до его конца, им предоставлялся выбор повернуть направо или налево. Справа их ожидало затемнение и пища, слева – удар слабым разрядом электрического тока. После серии таких «уроков» у червей вырабатывался рефлекс безошибочно направляться в нужную сторону, к пище.
Тип Кольчатые черви. Класс Малощетинковые
ОБЩАЯ ХАРАКТЕРИСТИКА
Строение и покровы . Тело малощетинковых червей (олигохет) сильно вытянутое, цилиндрическое, состоит из лежащих друг за другом колец , или сегментов . Все сегменты имеют сходное строение, т.е. для организации олигохет (и всех кольчатых червей) характерна повторяемость строения, или метамерия . Каждый сегмент, кроме самого первого, снабжен маленькими щетинками , обычно расположенными четырьмя пучками - парой боковых ипарой брюшных . Передний сегмент представляет собой головную лопасть - простомиум , лишенный глаз и антенн. Он несет ротовое отверстие . Последний сегмент - анальная лопасть , или пигидиум , - несет порошицу .
Покровы олигохет представлены эпителием , образующим на поверхности тонкую эластичную кутикулу (рис. 1). Эпителий богат железистыми клетками .
Кожно-мускульный мешок . Под эпителием расположены хорошо развитые мышечные слои . Наружный слой представлен кольцевыми мышечными волокнами . За счет сокращения данного слоя тело червя утончается и вытягивается. Внутренний слой , более мощный, представлен продольными мышечными волокнами , за счет сокращения которых тело червя утолщается и укорачивается.
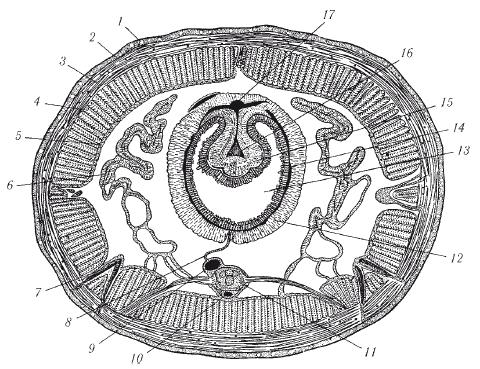
Рис. 1. Поперечный разрез средней части тела дождевого червя: 1 - кутикула; 2 - эпидермис; 3 - слой кольцевой мускулатуры; 4 - слой продольной мускулатуры; 5 - целомический эпителий; 6 - метанефридий; 7 - щетинка; 8 - мезентерий; 9 - брюшной сосуд; 10 - субневральный сосуд; 11 - брюшная нервная цепочка; 12 - хлорагогенные клетки; 13 - полость кишки; 14 - сосудистый плексус; 15 - тифлозоль; 16 - сосуд тифлозоля; 17 - спинной сосуд
Между кишечником и кожно-мускульным мешком находится вторичная полость тела , или целом -пространство, ограниченное собственными эпителиальными стенками мезодермального происхождения и содержащее целомическую жидкость (рис. 2). По строению целом отличается от первичной полости тела наличиемцеломической выстилки - собственной стенки. Выстилка образована двумя листками . Один прилегает к стенке тела, другой - к стенкам кишечника. Над кишечником и под ним оба листка срастаются, образуя брыжейку (мезентерий), которая делит целом на левую и правую стороны . Кроме этого, имеются поперечные перегородки , которые делят полость тела на камеры , соответствующие границам колец . Целом наполнен жидкостью, в которой плавают фагоциты, яйца, спермии. Целомическая жидкость, омывая внутренние органы, снабжает их кислородом и питательными веществами, а также способствует удалению продуктов обмена веществ и передвижению фагоцитов. Так же, как и жидкость, заполняющая первичную полость тела у круглых червей, целомическая жидкость может играть роль гидроскелета .
Пищеварительная система олигохет хорошо дифференцирована (рис. 3). Она начинается ротовым отверстием . Кишечник состоит из трех отделов - переднего , среднего и заднего . Наиболее дифференцирован передний отдел кишечника, состоящий из глотки, пищевода и мускульного желудка. Иногда перед желудком имеется зоб. В средней кишке для увеличения всасывательной поверхности образуется впячивание внутрь просвета кишечника - тифлозоль.

Рис. 2. Развитие целома у кольчатых червей. А-В - поперечные разрезы трех последовательных стадий развития сегмента: 1 - кишка; 2 - первичная полость тела; 3 - целом; 4 - наружная стенка целомического мешка; 5 - спинной мезентерий; 6 - внутренняя стенка целомического мешка; 7 - брюшной мезентерий; 8 - брюшные нервные стволы
Кровеносная система замкнутая и состоит из двух главных продольных сосудов - спинного и брюшного. Спинной сосуд проходит вдоль всего тела над кишкой, брюшной - под нею. Полость кровеносных сосудов представляет собой остатки первичной полости тела. Оба сосуда сообщаются кольцевыми сосудами, расположенными метамерно. Движение крови по сосудам обеспечивается пульсацией спинного сосуда и некоторых кольцевых сосудов передней части тела, называемых поэтому боковыми, или кольцевыми, сердцами. В спинном сосуде кровь течет вперед, в брюшном - назад. По кольцевым сосудам кровь движется из спинного сосуда в брюшной в передней части тела и в обратном направлении - в задних сегментах. Кровь может иметь красный цвет от железосодержащего близкого к гемоглобину позвоночных, растворенного в жидкости крови.

Рис. 3. Строение дождевого червя (вскрытый червь): 1 - глотка; 2 - пищевод; 3 - известковые железы; 4 - зоб; 5 - желудок; 6 - средняя кишка; 7 - спинная складка кишки (в разрезе); 8 - надглоточный нервный узел; 9 - узел брюшной нервной цепочки; 10 - спинной кровеносный сосуд; 11 - кольцевые сосуды, охватывающие глотку; 12 - брюшная нервная цепочка; 13 - метанефридии; 14 - семенники; 15 - семяпроводы; 16 - семенные мешки; 17 - яичники; 18 - яйцеводы; 19 - семяприемники; 20 - перегородки полости тела
Выделительная система представлена метанефридиями. Метанефридий начинается в полости тела воронкой - нефростомом. От воронки идет проток, который проходит через перегородку, входит в соседний сегмент и открывается наружу выделительной порой в боковой стенке тела. В каждом сегменте находится пара метанефридиев - правый и левый. Воронка и проток снабжены ресничками, вызывающими движение выделяемой жидкости.
Нервная система. Центральная часть нервной системы состоит из парных мозговых ганглиев - надглоточного и подглоточного, соединенных двумя окологлоточными коннективами (нервными стволами, соединяющими разноименные ганглии). Таким образом образуется окологлоточное кольцо. В центральную часть нервной системы входит и парный брюшной нервный ствол. В каждом сегменте стволы имеют утолщения - ганглии, которые соединяются между собой перемычками - комиссурами (поперечными нервными стволами, связывающими ганглии одного сегмента). Образуется брюшная нервная цепочка, похожая на лестницу. Каждый ганглий иннервирует все органы сегмента, в котором находится.
Органы чувств у олигохет очень слабо развиты в связи с роющим образом жизни. Глаза почти всегдаотсутствуют. Однако имеются светочувствительные клетки, в большом числе разбросанные в коже, что позволяет олигохетам иметь чувствительность к свету.
Половая система. Олигохеты - гермафродиты, но оплодотворение у них - перекрестное внутреннее. Половые гонады локализованы в половых сегментах. Мужские гонады - семенники - лежат в семенных капсулах, которые находятся в семенных мешках. Женская половая система представлена парой яичников, парой яйцеводов ияйцевыми мешками.
Размножение и развитие. Развитие прямое без стадии личинки. Яйца развиваются внутри яйцевого кокона, который формируется в районе пояска.
Помимо полового размножения у олигохет наблюдается бесполое размножение, похожее на бесполое размножение ресничных червей. Тело червя делится на две половины: у передней регенерирует задняя часть тела, у задней - головная часть тела.
Внутренности - viscera (splanchna). К ним относят органы пищеварения, дыхания, мочеотделения, размножения, расположенные в основном в грудной, брюшной и тазовой полостях (рис. 210).
![]()
Грудная полость - cavum thoracis расположена внутри грудной клетки. Полость выстлана внутригрудной фасцией - fascia endothoracia и серозной оболочкой - плеврой, сращенной с фасцией. В грудной полости различают три области: а) входную, краниальную на уровне первых двух костных сегментов грудной клетки; б) среднюю - от входа до уровня 7-го межреберья у крупного рогатого скота и лошадей и 6-го межреберья у свиней и собак; в) диафрагматическую - от вершины купола диафрагмы до прикрепления ее мышечного отдела к реберной стенке.
Брюшная полость - cavum abdominis краниально отграничена диафрагмой, каудально продолжается до входа в тазовую полость. Крышу брюшной полости составляют последние грудные и поясничные позвонки, расположенные под ними мышцы. Боковые и вентральные стенки образованы мышцами и фасциями живота. В краниальной части боковых стенок брюшной полости находятся последние ребра и вентрально-мечевидный хрящ.
Внутренняя поверхность брюшной полости выстлана поперечной фасцией - fascia transversalis и сращенным с нею париетальным (пристенным) листком серозной оболочки - брюшиной.
Брюшную полость условно разделяют на отделы и области. Различают три отдела (рис. 211):
1) краниальный - epigastrium (мысленно ограниченный сегментальной плоскостью, поставленной касательно каудальному краю последних ребер. Краниальнее этой плоскости до диафрагмы и располагается этот отдел). В отделе разграничивают три области (рис. 211): вентрально лежит область мечевидного хряща - regio xiphoidea, ее дорсальная граница проходит по линиям реберных дуг; дорсально между реберной стенкой и диафрагмой и по бокам расположены правое и левое подреберье - regio hypochondriaca dextra et sinistra;

2) средний брюшной отдел - mesogastrium расположен между передним отделом и плоскостью, перпендикулярной позвоночнику на уровне маклоков. В среднем отделе четыре области: правая и левая подвздошные области - regio iliaca dextra et sinistra лежат между боковыми стенками брюшной полости до парасагиттальных плоскостей, проведенных мысленно на уровне концов поперечных отростков поясничных позвонков; пупочная область - regio umbilicalis - в вентральной части средней брюшной полости; поясничная область - regio lumbalis, или почечная область, располагается дорсально над мысленно проведенной плоскостью на уровне середины последнего ребра;
3) задний брюшной отдел -- hypogastrium расположен от сегментальной плоскости на уровне маклоков до входа в тазовую полость. В этом отделе три области: правая и левая паховые - regio inguinalis dextra et sinistra являются продолжением подвздошных областей; между ними находится лонная область (срамная) - regio pubis - является продолжением пупочной области.
Тазовая полость - cavum pelvis. Стенки тазовой полости образованы костями тазового пояса, крестцовой костью, первыми хвостовыми позвонками, крестцово-седалищными связками и мышцами. Полость выстлана подвздошной и тазовой фасциями. В тазовой полости различают: а) краниальный вход - apertura pelvis cranialis, обозначенный пограничной линией, проходящей по мысу, затем краниальному краю крыльев крестцовой кости, подвздошно-лонному гребню до лонного бугорка; б) каудальный вход (выход) - apertura pelvis caudalis, ограниченный вентрально-седалищной дугой, по бокам крестцово-седалищными связками (правой и левой), дорсально первыми хвостовыми позвонками.
Между краниальным и каудальным входом можно выделить среднюю часть тазовой полости. В средней части находится дно костного таза в виде углубления между правыми и левыми лонными и седалищными костями.
Псевдоцель (рис. 4.21, Б) у нематод и коловраток является остатком полости бластулы, возникающей на ранних этапах эмбриогенеза. Снаружи он ограничен эктодермой, а изнутри - энтодермальной стенкой пищеварительного тракта. Внутренние органы расположены в первичной полости свободно, и пространство между ними заполнено крупными вакуолизированными мезодермальными клетками. Таким образом, органы пространственно разделены и функционируют независимо друг от друга. Животное может быстро менять форму тела благодаря тому, что клетки мезодермы легко деформируются. Псевдоцель как бы предшествует появлению внутренней полости другого типа - целома.
Целом мы называем вторичной полостью тела , поскольку он появляется в процессе эмбриогенеза после бластоцеля. По мере развития целома бластоцель редуцируется, превращаясь в ряд заполненных кровью пространств, ограниченных мезодермальными стенками. Целом образуется как щель между клетками мезодермы и выстлан соответственно клетками мезодермального происхождения, образующими брюшину . Участок брюшины, расположенный вертикально, получил название брыжейка . Пищеварительный тракт, отделенный от стенок тела, оказывается подвешенным на брыжейке. Целом заполнен целомической жидкостью .
По данным эмбриологических исследований целом у разных животных возникает по-разному, и пока еще не ясно, какой из способов образования целома наиболее древний. На рис. 4.22 показаны три способа образования целома.
Рис. 4.22. Способы образования целома. Схизоцель характерен для моллюсков, членистоногих, кольчатых червей, продвинутых хордовых; целомическая полость возникает в результате образования щели в мезодерме зародыша. Энтероцель характерен для иглокожих, полухордовых, головохордовых, примитивных хордовых (мезодерма отделяется от энтодермы в результате образования выростов кишки). Гоноцель характерен для кольчатых червей (посегментно расположенные полости гонад вместе составляют постоянную полость - целом; смежные передние и задние стенки соседних полостей образуют перегородки, разделяющие сегменты у кольчатых червей). Мезодерма окружает архентерон (первичную полость тела)
В результате развития целома слой мезодермы, прилегающий к эктодерме (соматический слой , или соматоплевра ), входит в состав стенки тела, а мезодермальный слой (внутренностный слой , или спланхноплевра ), прилегающий к энтодерме пищеварительного тракта, образует мускулатуру кишечника. Относительно неподвижная целомическая жидкость отделяет пищеварительный тракт от стенок тела, а органы, которые выдаются в полость тела, удерживаются брюшиной.
70. ^ Вторичная полость тела, её функции и развитие.
У плоских червей все пространство м/у эктодермой и энтодермой занимает мезодерма, образующая плотный промежуточный слой. Единственная полость тела представлена первичной кишкой , / архентероном . Но поскольку эта полость на самом деле сообщаться с окружающей средой, ее нельзя рассматривать как истинную внутреннюю полость тела (целом). Строение животных, подобное этому, следует определить как ацеломическое . У всех высших мн/кл животных формируется обширная внутренняя полость тела – целом . Морфологически целом отличается от первичной полости тела наличием собственных клеточных стенок, кот. в онтогенезе всегда формируются за счет мезодермы. Стенки, ограничивающие целом, представляют собой 1слойный эпителий, кот. наз. целомическим / перитонеальным эпителием . Этот эпителий одевает внутреннюю поверхность стенки тела, прилегает к кишечнику и ко всем внутренним органам. За его счет образуются особые каналы, сообщающие полость целома с внешней средой. Т.о., целом представляет собой не просто пространство м/у внутр. органами, но вполне оформленный орган. Целом не сплошной, он поделен продольно на 2 половины, правую и левую, двухслойной клеточной перегородкой. Она начинается как бы в виде складки перитонеального эпителия по срединной спинной линии, спускается книзу, слои ее охватывают с двух сторон кишечник, а затем под ним вновь соединяются и, достигая брюшной стенки тела, переходят опять в перитонеальный эпителий стенки тела. Продольной перегородкой – брыжейкой кишечник подвешен к стенкам тела. На границе м/у каждыми двумя сегментами перитонеальный эпителий образует двухслойную перегородку – септу , пересекающую полость тела. Септами целом делится на известное число поперечных участков, обычно соответствующих числу сегментов тела. В нек. отделах септы м. ин. редуцироваться. Целом заполнен водянистой жидкостью, в кот. плавают амебоидные кл. Функции: 1)является гидростатическим скелетом; 2)позволяет пищеварительному тракту и стенка тела функционировать независимо др. от др.; 3)делает возможным значительное размеров тела; 4)обеспечивает транспорт пит. в-в, конечных продуктов обмена и газов, кот. осуществляется с помощью жидкости, заполняющей целом; 5) временно может накапливать конечные продукты обмена и избыток жидкости; 6)позволяет размеры внутренних органов; 7) участвует в осморегуляторных процессах в организме; 8)созревание половых клеток – спермиев и яиц.
РАЗВИТИЕ. Псевдоцель у нематод и коловраток явл. остатком полости бластулы, возникающей на ранних этапах эмбриогенеза. Снаружи он ограничен эктодермой, изнутри – эндодермальной ст. ПТ. Внутренние органы расположены в первичной полости свободно, и пространство м/у ними заполнено крупными вакуолизированными мезодермальными кл. Т.о. органы пространственно разделены и ф-ют независимо др. от др. Животное м. быстро менять форму тела due to, что кл. мезодермы легко деформируются. Псевдоцель предшествует появлению целома. Целом – это вторичная полость тела, поскольку он появляется в эмбриогенезе после бластоцеля. По мере развития целома бластоцель редуцируется, превращаясь в ряд заполненных кровью пространств, ограниченных мезодермальными стенками. В результате разв. целома слой мезодермы, прилегающий к эктодерме, входит в состав стенки тела, а прилегающий к энтодерме ПТ, обр. мускулатуру кишечника. У членистоногих и моллюсков полость тела наз. гемоцелем . Это – сильно бластоцель, почти полностью вытесняющий целом. Бластоцель представлен синусами, заполненными кровью (гемолимфа), кот. обычно циркулирует по гемоцелю. Стенки гемоцеля никогда не уч. в обр. гонад. Целом сохраняется только в выделительных органах и протоках половых желез. По Догелю : полость тела всех членистоногих им. двойственное происхождение. Во время эмбриогенеза у них в > случаев закладывается сегментированный целом, а впоследствии стенки целомических мешков разрушаются и целомические полости сливаются как др. с др., так и с остатками первичной полости тела, т.о. формируется смешанная полость тела / миксоцель , в кот. располагаются внутренние органы.
^ СПОСОБЫ ОБРАЗОВАНИЯ ЦЕЛОМА :
Схизоцель (моллюски, членистоногие, кольчатые черви, продвинутые хордовые) – целомическая полость возникает в результате образования щели в мезодерме зародыша.
Энтероцель (иглокожие, полухордовые, головохордовые, примитивные хордовые) – мезодерма отделяется от энтодермы в результате образования выростов кишки.
Гоноцель (кольчатые черви) – посегментно расположенные полости гонад вместе составляют постоянную полость – целом; смежные передние и задние стенки соседних полостей обр. перегородки, разделяющие сегменты у кольчатых червей; мезодерма окружает архентерон (первичную полость тела).
Целом – вторичная полость тела у всех животных, которым свойственен целом.
При энетероцельном происхождении мезодермальный целом, мешок образуется путем изменения, выпячивания первичногокишечника.
При телобластическом, когда образуются телобластический тяжи, внутри них появляется цель, которая превращается в целом.
Целомический мешок формируется симметрично по бокам кишечника. Стенка каждого целомического мешка образванного к кишечнику называется стианоплеврой, а в сторону к эктодерме называется соматоплеврой. Т.о. в ходе развития зародыша формируются различные полости, имеющее важное морфогенетическое и функциональное значение. Сначала появляется полость Бэра, затем она превращается в первичную полость тела (бластоцель), далее в процессе гаструляции образуется гастроцель, а затем целом.
При образовании гастроцели и целома бластоцель уменьшается и от нее остаются щели в промежутках между стенками кишечника и целома. Эти щели превращаются в полости кровеносной системы. Гастроцель превращается в полость средней кишки.
Три способа образования целома.
спизоцель – характерна для моллюсков, кольчатых червей – целомная полость возникает в результате образования щели в мезодерме зародыша
эктероцель – характерна для иглокожих, полухордовых. Мезодерма отделяется от эктодермы в результате образуется вырост клетки.
Гоноцель – характерен для кольчатых червей
Возможность значительно увеличить размеры тела
обеспечивает транспорт питательных веществ, конечных продуктов обмена и газов, которые с током жидкости попадают в полость тела.
временно могут накапливать продукты обмена и избыток жидкости
позволяет увеличить размеры внутренних органов
участвует в осморегуляционных процессах в организме.
71. Механизмы возбуждения. Проведение возбуждения, синоптическая передача.Возбуждение – активная реакция специализированных (возбудимых) кл. на внешнее воздействие, проявляющаяся в том, что кл. нач. вып. присущие ей специф. ф-ии. Возбудимая кл. м. нах. в 2 сост: покоя и возбуждения . 3 типа возбудимых кл. : Н (генерация эл. импульса); мышечные (сокращение); секреторные (выброс в межкл. пр-во БАВ). М возбудимой кл поляризована, т.е. есть пост. разность потенциалов м/у внутр. и нар. пов-тью М, кот. наз. мембранный потенциал (МП) . В состоянии покоя МП -60..-90 мВ (внутр. ст. М (-) к нар). Значение МП кл. в сост. покоя - потенциал покоя (ПП). ↓ МП относительно ПП наз. деполяризацией , а - гиперполяризацией. Реполяризация – восст. исх. ур. МП. При действии слабых (подпороговых) импульсов эл. тока в кл. развив. электротонический потенциал (ЭП) – сдвиг МП кл., вызываемый действием пост. эл. тока. ЭП - пассивная реакция кл. на эл. раздражитель; состояние ионных каналов и транспорт ионов при этом не изменяется. ЭП не проявляется физиол. реакцией кл. Поэтому ЭП не явл. возбуждением. При действии > сильного подпорогового тока возникает > длинный сдвиг МП – локальный ответ (ЛО) – актив. р-ия кл. на эл. раздр-тель, но, состояние ионных каналов и транспорт ионов при этом изменяется незначительно. Не проявл. заметной ФРК. ЛО наз. местным возбуждением , При действии порогового и сверх- тока в кл. разв. потенциал действия (ПД) - значение МП кл. оч. быстро ↓ до 0 (деполяризация), а затем МП становится (+) (+20..+30 мВ), т.е. внутр. ст. М (+) к внешней ст. Затем значение МП быстро возвр. к исх. ур. ПД - распространяющееся возбуждение , т.к., возникнув в 1 уч. М, он быстро распр. во все стороны. ^ РАЗВИТИЕ ПД : Деполяризация . Развитие ПД возможно только при действии раздражителей, кот. вызыв. деполяр. кл. М, если до крит. ур. деполяр. (КУД) происх. лавинообразное открытие потенциалчувств. Na + -каналов. Ионы Na + вх. в кл. по градиенту [с] (натриевый ток) → МП оч. быстро ↓ до 0, а затем приобретает (+) значение. Явление изменения знака МП называют реверсией заряда М. Быстр. и медл. реполяризации . В результате деполяризации М происходит открытие потенциалчувств. К + -каналов. Ионы К + вых. из кл. по градиенту [с] (калиевый ток) → восст. МП. В начале фазы интенсивность калиевого тока и реполяризация происходит быстро, к концу фазы интенсивность калиевого тока ↓и реполяризация замедляется. Гиперполяризация разв. за счет остаточного К тока и за счет прямого электрогенного эффекта активировавшейся Na + /K + помпы. Овершут – Т t, в теч. кот. МП им. (+) значение. Пороговый потенциал – разность м/у МП покоя и КУД. Чем он >, тем Супернормальная возбудимость – возбудимость нормальной. Во t нач. деполяризации и во t медленной реполяризации. возбудимости кл. в эти фазы ПД обусловлено ↓ порогового потенциала по сравнению с нормой. Абсолютная рефрактерность – возбудимость падает до 0. Во t деполяризации кл. невозбудима, ибо все ее Na + -каналы уже нах. в откр. сост. Относительная рефрактерность – возбудимость кл. значительно ↓ нормальной, только оч. сильные раздражители м. вызвать ее возбуждение. Во t реполяризации каналы возвращаются в закрытое сост. и возбудимость кл. постепенно восст. Субнормальная возбудимость - незначительное ↓ возбудимости кл. ниже нормального уровня. Это происх. из-за порогового потенциала во t гиперполяризации. ЗАКОНЫ: Анатом. и физиол. непрерывн., двустороннего проведения возбужд. и изолир-ого проведения – си распростр. по волокну, вх. в состав Н, то не перед. на сосед Н волокна. ЛО (местное возбуждение) – распростр. по Н волокнам с затуханием, т.е. амплитуда ЛО быстро ↓ с расстояния от места его возник; вследствие затухания ЛО распростр. на небольшие расстояния (не > 2 см); местное возбуждение распространяется пассивно, без затрат Е кл; такой способ распростр. возбуждения называют электротоническим. ПД (распространяющееся) – распростр. по Н волокнам без затухания, амплитуда ПД одинакова на любом расстоянии от места его возник; расстояние, на кот. распростр. ПД, ограничено тока длиной Н волокна; распростр. ПД – активный пр-с, в ходе кот. изм. состояние ионных каналов волокна, Е АТФ требуется для восстановления трансМ ионных градиентов; По немиелиновым . В состоянии покоя вся внутр. пов-ть М Н волокна несет (-) заряд, а нар. ст. М – (+). Эл. ток м/у внутр. и нар. ст. М не протекает, т.к. М им. высокое эл. сопротивление.Во t развития ПД в возбужденном уч. М происх. реверсия заряда. На границе возбужденного и не- уч. нач. протек. эл. ток, кот раздраж. ближ. уч. М и → его в сост. возбужд, в то t как ранее возбужд. уч. возвр. в сост. покоя. Охватываются новые уч. М Н волокна. По миелиновым . В Н волокне уч. М, покрытые миел. оболоч., явл. невозбуд.; возбуждение м. возникать только на уч. М в обл. перехватов. Т.о. возбуждение распростр. по М скачкообразно от 1 перехвата к др.
72. ^ Выделительная система. Функция почек. Почки – гл. орган выделения и осморегуляции. Функции почек:
1) выведение из организма воды, аммиака, солей, мочевины, мочевой кислоты;
2) del чужерод. и яд. в-в, образующихся в организме / принятых в виде лекарств, при курении и т.д;
3) поддерж. постоянной кислотно-щелочной реакции крови. При накоплении в крови кислых / щелочных продуктов обмена ч/з почки увеличивается выведение излишков соответствующей солей. Например, при питании мясной пищей в организме образуется много кислых продуктов обмена, соответственно и моча становится кислой; - растительной пищи реакция мочи становится щелочной;
4) регуляция хим. состава жидкостей тела путем удаления в-в, кол-во кот. текущую потребность;
5) регуляция содержания воды в жидкостях тела (и тем самым их V) и рН этих жидкостей;
6) соли, в состав кот. вх. аммиак, выводятся из организма, а Na и K сохраняются для нужд организма;
По обе стороны тела на ур. грудных и поясничных позвонков. Вес 1 почки 0,5% общего веса тела; левая почка слегка вытянута по ср. с пр. Кровь поступает в почки ч/з почечные артерии, отходящие от аорты, а оттекает от них почечными венами, впадающими в нижнюю полую вену. Обр. в почках моча идет по мочеточникам в мочевой пузырь, где накапливается до выделения ч/з мочеиспускат. канал. На поперечном разрезе: ближе к пов-сти корковое в-во и внутреннее мозговое вещество . Корковое в-во покрыто фиброзной капсулой и сод. едва заметные клубочки . Мозговое в-во сост. из канальцев, собирательных трубок и кровеносных сосудов, собранных вместе в в. почечных пирамид . Верхушки пирамид, наз. сосочками , открываются в почечную лоханку , обр. расширенное устье мочеточника. Ч/з почки проходит множество сосудов, обр. густую сеть капилляров. Осн. стр-ой и функц. единичей почки явл. нефрон вместе с его кровеносными сосудами. У чела в 1 почке ок. 1 млн нефронов по 3 см к. → огромная пов-сть для обмена в-ми. К. нефрон вкл. 6 отделов , с. различ. по строению и физиол. функ-ям:
1 ) почечное тельце (мальпигиево тельце),
) почечное тельце (мальпигиево тельце),
сост. из боуменовой капсулы и клубочка;
2) проксимальный извитой каналец;
3) нисходящее колено петли Генле;
4) восходящее колено петли Генле;
5) дистальный извитой каналец;
6) собирательная трубка.
^
Корковые нефроны
расположены в коре и им. относительно кор. петли Генле, кот. лишь недалеко заходят в мозговое в-во. В юкстамедуллярных
нефронах почечные тельца расположены около границы коры и мозгового слоя (от лат. juxta = рядом). Они им.длин. нисх. и восх. колена петли Генле, глубоко проник. в мозговое в-во. При нормальном кол-ве воды в орг-ме V плазмы контролируют корковые нефроны, а при недостатке воды происходит ее усиленная реабсорбция в юкстамодуллярных нефронах. 1) Ультрафильтрация
. В клубочке все низкомол. в-ва, такие как, клюю, вода, мочевина, переходят в фильтрат, заполняющий боуменову капсулу и поступающий затем в каналец нефрона; 2) Избират. реабсорбция
. Все в-ва, кот. м. б. исп. орг-ом / нужны для поддержания водно-солевого состава жид-тей тела, всас. из фильтрата в капилляры (глю всас. в проксимальном извитом канальце); 3) Секреция
. До того как фильтрат покинет нефрон в в. мочи, в него м. секретироваться ненужные орг-му в-ва; напр., избыток ионов K, H и NH 4 секретируются кл. дистального извитого канальца. 1. Активный транспорт
. В 2) и 3) мол. и ионы активно секрет. в фильтрат и всасываются из него (глю в капилляры из проксимального и NaCl – в толстом восходящем колене). 2. Избирательная проницаемость
. Разл. уч. нефрона обл. избир. прониц. для ионов, воды и мочевины. Проксимал. относит. мало прониц. по ср. с дистальными канальцами. Проницаемость собир. трубки м. регулироваться гормонами. 3. Концентрационные градиенты
. В результате дей-ия 1) и 2) в тканевой жидкости мозгового в-ва поддерж. концентр. градиенты. 4. Пассивная диффузия и осмос
. Ионы Na, Cl и мочевина б. диффундировать в фильтрат и из него по [с] градиенту в тех уч. нефрона, кот. проницаемы для них. Вода путем осмоса в тканевую жид-сть там, где она б. гипертонична. 5. Гормональная регуляция
. Гормоны АДГ, альдостерон и др. дей-ют на дистальные извит. канальца и собир. трубку.
73
.Механизмы мышечного сокращения. Регуляция мышечного тонуса и движений.
^
С труктура миофибрилла и ее изменения при сокращении.
В П-П мышечных волокнах миофибриллы разделены на правильно чередующиеся участки (диски), обладающие разными оптическими св-ми. Анизотропные
(А) участки обл. двойным лучепреломлением (в обыкн. свете они выглядят темными) и изотропные
(I) (выглядят белыми). Посередине диска А различается светлая полоска Н
, поседерине диска I – темная полоска Z
, представляющая собой тонкую М, сквозь поры кот. проходят миофибриллы. Каждая миофибрилла сост. в ср. из 2500 протофибрилл
, представляющих собой удлиненные полимеризованные молекулы миозина
и актина
. Миозиновые нити в 2е толще актиновых. В состоянии покоя
нити расположены в миофибрилле т.о., что тонкие длинные актиновые нити вх. св. концами в промежутки м/у толстыми и > короткими миозиновыми нитями. Благодаря этому диски I состоят только их актиновых нитей, а диски А – из нитей миозина. Светлая полоска Н представляет собой узкую зону, свободную от актиновых нитей. М Z, проходя ч/з середину диска I, скрепляет м/у собой эти нити. Многочисленные поперечные мостики соединяют м/у собой Миозиновые и актиновые нити. При сокращении
указанные нити не укорачиваются, в начинают скользить др. по др: актиновые нити вдвигаются в промжутки м/у миозиновыми, в результате чего диски I укорачиваются, а диски А сохраняют свой размер. Почти исчезает светлая полоска Н, т.к. актиновые нити при скольжении сближаются др. с др. св. концами. СОКРАЩЕНИЕ
. Инициатором сокращения скелетной мышцы явл. ПД, распространяющийся при возбуждении вдоль пов-сти М мышечного волокна. Важным промежуточным звеном м/у деполяризацией М и началом мышечного сокращения явл. проникновение в область миофибрилл свободных ионов Са, дей-ие кот. опосредуется комплексом Б, тесно св. с актин. филаментами. Один из них - тропомиозин
- жесткая стержневидная мол., св. с актин. филаментом по всей его длине и придающая ему жесткость. Др. – тропонин –
это комплекс из 3 ПП - тропонинов T, I
и C
. Тр. Т св. с тропомиозином и опр. положение всего комплекса на тонком филаменте. Тр. I отвечает за ингибирование присоед. миозиновых поперечных мостиков к актин. филаменту. Ингибиторный эффект тр. I - стерическое препятствие для такого взаимодействия. Тр. С явл. центром св. ионов Са. В сост. покоя тропониновый комплекс размещен так, что тропомиозин и тр. I инг. присоед. мио к актин. [Са] в цитизоле приводит к его св. с тр. С → меняется конфор-ция всего комплекса, происх. смещение тр. I, откр. миозинсв-щий центр. Пока ионы Са не будут удалены из тропонинового комплекса. Одиночный ПД вызывает [Са] всего на 30 мс, этого достаточно только для 1 взаимодействия миозиновых головок с актином (это min мышечная сила). Если ПД в мышечном волокне возникают с интервалами саркоплазматический ретикулум. СР обволакивает наподобие полой манжеты каждую миофибриллу. На уровне Z-пластины к. саркомера цистерны СР вступают в тесный контакт с Т-трубочкой и как бы сжимают ее м/у собой. Т-трубочки анатомически соед. пов-ную М с миофибриллами, лежащими в глубине мышечного волокна и обеспечивают перенос ПД внутрь к к. миофибрилле и цистерн СР. Эл. возбуждение Т-трубочек приводит к открытию в М цистерн СР Са каналов, и ионы Са выходят в цитозоль где находятся миофибриллы. Именно этот внезапный подъем [с] свободных ионов Са вызывает их сокращение. [Са] кратковременно, поскольку ионы Са быстро перекачиваются обратно в цистерны СР Са-АТФ-азой. Обычно возвращение [Са] в цитозоле к ур. контроля происходит за 30 мс, что приводит к расслаблению миофибрил.
труктура миофибрилла и ее изменения при сокращении.
В П-П мышечных волокнах миофибриллы разделены на правильно чередующиеся участки (диски), обладающие разными оптическими св-ми. Анизотропные
(А) участки обл. двойным лучепреломлением (в обыкн. свете они выглядят темными) и изотропные
(I) (выглядят белыми). Посередине диска А различается светлая полоска Н
, поседерине диска I – темная полоска Z
, представляющая собой тонкую М, сквозь поры кот. проходят миофибриллы. Каждая миофибрилла сост. в ср. из 2500 протофибрилл
, представляющих собой удлиненные полимеризованные молекулы миозина
и актина
. Миозиновые нити в 2е толще актиновых. В состоянии покоя
нити расположены в миофибрилле т.о., что тонкие длинные актиновые нити вх. св. концами в промежутки м/у толстыми и > короткими миозиновыми нитями. Благодаря этому диски I состоят только их актиновых нитей, а диски А – из нитей миозина. Светлая полоска Н представляет собой узкую зону, свободную от актиновых нитей. М Z, проходя ч/з середину диска I, скрепляет м/у собой эти нити. Многочисленные поперечные мостики соединяют м/у собой Миозиновые и актиновые нити. При сокращении
указанные нити не укорачиваются, в начинают скользить др. по др: актиновые нити вдвигаются в промжутки м/у миозиновыми, в результате чего диски I укорачиваются, а диски А сохраняют свой размер. Почти исчезает светлая полоска Н, т.к. актиновые нити при скольжении сближаются др. с др. св. концами. СОКРАЩЕНИЕ
. Инициатором сокращения скелетной мышцы явл. ПД, распространяющийся при возбуждении вдоль пов-сти М мышечного волокна. Важным промежуточным звеном м/у деполяризацией М и началом мышечного сокращения явл. проникновение в область миофибрилл свободных ионов Са, дей-ие кот. опосредуется комплексом Б, тесно св. с актин. филаментами. Один из них - тропомиозин
- жесткая стержневидная мол., св. с актин. филаментом по всей его длине и придающая ему жесткость. Др. – тропонин –
это комплекс из 3 ПП - тропонинов T, I
и C
. Тр. Т св. с тропомиозином и опр. положение всего комплекса на тонком филаменте. Тр. I отвечает за ингибирование присоед. миозиновых поперечных мостиков к актин. филаменту. Ингибиторный эффект тр. I - стерическое препятствие для такого взаимодействия. Тр. С явл. центром св. ионов Са. В сост. покоя тропониновый комплекс размещен так, что тропомиозин и тр. I инг. присоед. мио к актин. [Са] в цитизоле приводит к его св. с тр. С → меняется конфор-ция всего комплекса, происх. смещение тр. I, откр. миозинсв-щий центр. Пока ионы Са не будут удалены из тропонинового комплекса. Одиночный ПД вызывает [Са] всего на 30 мс, этого достаточно только для 1 взаимодействия миозиновых головок с актином (это min мышечная сила). Если ПД в мышечном волокне возникают с интервалами саркоплазматический ретикулум. СР обволакивает наподобие полой манжеты каждую миофибриллу. На уровне Z-пластины к. саркомера цистерны СР вступают в тесный контакт с Т-трубочкой и как бы сжимают ее м/у собой. Т-трубочки анатомически соед. пов-ную М с миофибриллами, лежащими в глубине мышечного волокна и обеспечивают перенос ПД внутрь к к. миофибрилле и цистерн СР. Эл. возбуждение Т-трубочек приводит к открытию в М цистерн СР Са каналов, и ионы Са выходят в цитозоль где находятся миофибриллы. Именно этот внезапный подъем [с] свободных ионов Са вызывает их сокращение. [Са] кратковременно, поскольку ионы Са быстро перекачиваются обратно в цистерны СР Са-АТФ-азой. Обычно возвращение [Са] в цитозоле к ур. контроля происходит за 30 мс, что приводит к расслаблению миофибрил.
74. Эндокринная система и её регуляторные функции.
Основная железа внутренней секреции - гипофиз , кот. сост. из задней доли (нейрогипофиз ) и передней доли (аденогипофиза ). Гипофиз выделяет по меньшей мере 9 гормонов, многие из которых регулируют активность др. эндокринных желез. Сам гипофиз нах. под контролем гипоталамуса. Гормоны нейрогипофиза . В нейрогипофизе секретирует два важнейших гормона: окситоцин и вазопрессин , кот. обр. в рез. гидролиза общего предшественника, вырабатываемого в гипоталамусе. Окситоцин регулирует тонус гладкой мускулатуры, особенно мускулатуру матки беременных и по этой причине используется в качестве фармакологического средства для стимуляции родов. Вазопрессин кровяное давление, действуя постепенно и длительно. Этот гормон оказывает сильное влияние на водно-солевой обмен, ↓ диурез (выделение мочи). Поэтому вазопрессин наз. антидиуретическим гормоном (АДГ). При его недостатке выделение мочи и развивается несахарный диабет .
^ Гормоны аденогипофиза . В аденогипофизе образуются гормоны регулирующие активность др. эндокринных желез и гормоны, действующие непосредственно на ткани мишени. К первой группе относятся: 1)адренокортикотропный гормон (АКТГ) , стимулирующий синтез и секрецию стероидных гормонов корой надпочечников; 2)тиреотропный гормон , усиливает синтез и секрецию тиреоидных гормонов щитовидной железой; 3)гонадотропные гормоны , действуют на половые железы. Гормоны аденогипофиза, действующие на ткани-мишени непосредственно: 1)соматотропный гормон (СТГ) / гормон роста , стимулирующий синтез РНК и белка и рост всех тк; 2)пролактин - стимулирует синтез белка молока и развитие молочных желез. Надпочечники сост. из 2 типов железистой ткани: нар. коры и внутр. мозгового слоя. На кору надпочечников воздействует АКТГ, который стимулирует в ней синтез из холестерола и секрецию целой группы кортикостероидов. В эту группу входят: минералокортикоиды , регул. функцию почек, и глюкокортикоиды . У чела осн. глюкокортикоид - кортизол . В течение суток концентрация кортизола подчиняется суточному ритму. Кортизол и др. глюкокортикоиды обладают шир. спектром действия, в том числе и противовоспалительным. Но наиболее важный эффект глюкокортикоидов заключается в стимуляции в печени глюконеогеза . В процессе глюконеогеза активируется катаболизм мышечных белков, а образующиеся аминокислоты метаболизируются с образованием глюкозы. В физиологических условиях все эффекты глюкокортикоидов направлены на быстрое обеспечение организма Е (особенно при стрессах). При стрессе глюкокортикоиды + играют пермиссивную роль в действии катехоламинов на гладкую мускулатуру сосудов. Катехоламины в мозговом слое . При стрессе они синтезируются и выделяются в кровь в большом количестве, что → к усилению сердечной деятельности, + они сокращают гладкую мускулатуру и сужают просвет КС в разл. орг. и тк. (кр. скелетной мускулатуры) → происходит перераспределение кровотока т.о., что улучшается снабжение мышц О 2 и пит. в-ми (утекай!). А только в присутствии кортизола. Действуют разнонаправлено: адреналин частоту сердечных сокращений и ↓ общее сопротивление периферических сосудов, а нор – наоборот, ибо адреналин св. с β-адренэргическими Р → активируется АЦ, а нор с α-адренэргическими Р → АЦ инг. и внутрикл. ур. Са 2+ . Они сужают сосуды кожи и кровяное давление. Щитовидная железа выделяет тиреоидные гормоны (L-тироксин и трииодтиронин ), кот. воздействуют на печень, почки, сердце, НС и скелетные мышцы, стимулируя кл. дых. и метаболизм → теплопродукции - важная роль в терморегуляции. Совместно с СТГ, тиреоидные гормоны стимул. синтез Б в период разв. Сод. йод (суточное 150 мкг), есть запас на неск. мес., потом гипотиреоз . Гипотиреоз → к ↓ интенсивности всех метабол. пр-сов → к замедленной реакции на разл. внешние раздражители и утомляемости. В островках Лангерганса поджелудочной железы - глюкагон и инсулин . Инсулин оказывает регулирующее действие на метабол. пр-сы + стимул. поглощ. глю почти всеми кл. (кроме Н), облегчая прохождение ее ч/з М. В жир. и мыш. кл. им. ЦП визикулы, сод. Б - трансМ переносчики глю. Повышенный уровень инсулина в крови и связывание его с поверхностными инсулиновыми рецепторами, является сигналом для начала движения д. визикул к ПМ, выхода из них Б-транспортеров, встраивания их в М и резкого поглощения глю кл, в 1 очередь жир. и мыш. В свою очередь синтез и выделение инсулина регулируется ур. глю в крови. При кот. выделение инсулина , ↓ же → к прекращению выделения гормона и к удалению из ЦПМ Б-транспортеров глю. Суммарное действие инсулина → к ↓ [глю] в кр. (0,8-1,0 г/л). Так как ЦНС получает всю необходимую для ее А Е в результате окисления глю, то ↓ ур. глю в крови до 0,2-0,5 г/л → к гипогликемическому шоку (помутнение сознания, кома). Глюкагон явл. антагонистом инсулина.
75. Ранние ст. зародыш. развития (дробление, гаструляция, нейруляция). Органогенез.
У всех жив. З. р. складывается из оплодотворения / (при партеногенезе) активации яйца, делений дробления, гаструляции, органогенеза и выхода из оболочек или рождения.
Начало З. р. - оплодотворение - происходит в материнском организме / в водной среде. Подвижный сперматозоид достигает яйца и проникает в него, часто ч/з специальные отверстия в оболочках - микропиле . Биосмысл сост. в обмене генет. инфой м/у жив. 1 популяции, т.к. каждый новый организм сочетает в себе наследств. признаки 2 родителей. 1n + 1n = 2n.
Дробление - это многокр. митозы зиготы, в результате кот. зародыш становится многокл., не меняя при этом существенно своего V. Образование многоклеточности 1 и осн. биороль Д. 2ая роль сост. в восст. ядерно-плазматического соотношения, кот. ↓ в ходе ст. большого роста ооцита. Видовые особенности процесса дробления определяются двумя основными параметрами: 1) кол-вом и распр. желточных Б в ЦП (желток подавляет дробление); 2) присутствием в ЦП факторов, кот. влияют на ориентацию митотического веретена и время его образования.
^ 1) Гомолецитальные яйца (сод. сравн. мало равномерно распр. Ж) претерп. полное равномерное Д . Чаще Ж распр. в ЦП яйца неравномерно (телолецитальные и центролецитальные ). Обл., сод. > Ж, делится медленнее бедной Ж → полное неравномерное Д ., / совсем не делится → частичное Д. Яйца, с полным Д. – голобластические (гомолецитальные млекопит., ланцетника; и часть телолецитальных > земноводных, кот. им. полное, но неравномерное Д.), с частичным - меробластические (часть телолецитальных и центролецитальные с большим кол-вом Ж у > членистоногих). В таких телолецитальных яйцах дел. только бедная Ж анимальная часть, кот. разделяется на 2, 4 и > число бластомеров, обр. диск кл. на пов-ти недробящегося Ж, - дискоидальное Д . (рыбы, птицы). В результате дискоидального Д. обр. дискобластула . После нескольких синхронных делений, ядра с окружающей их ЦП, перемещаются по ЦП мостикам в поверхн. слой ЦП, кот. сначала предст. с. симпласт , затем в/г к. Я обособляется отдельная кл. → обр. зародыш, ст. кот. сост. из 1 сл. кл. (бластодермы), а центр. часть занята неразделившимся Ж с находящимися в нём кл; такой зародыш наз. перибластулой , а Д. - поверхностным, / синцитиальным .
2) Т.к. плоскость дробления всегда перпендикул. оси веретена деления, то положение веретён опред. положение бластомеров др. относ. др. По этому признаку при полном Д. различают радиальное , спиральное , билатеральное и двусимметричное Д . При радиальном Д. (земноводные) бластомеры располагаются так, что любая плоскость, кот. м. провести ч/з анимальную ось яйца, явл. плоскостью симметрии. Первые 2 борозды проходят меридионально, а 3-я - экваториально; затем происх. их чередование. В итоге обр. многокл. пузырёк с полостью - целобластула . При спиральном Д. (моллюски) микромеры, отделяющиеся от первых 4 бластомеров (макромеров), распол. в промежутках м/у ними. Происх. смещения бластомеров верхнего яруса относ. нижнего вправо - дексиотропное Д . / влево - леотропное Д . Зародыш на ст. бластулы им. полость (неравномерная целобластула ) / не им. её (стерробластула ). При билатеральном Д. (круглые червей), а также на поздних ст. спирального Д. деления происх. так, что у зародышей им. только 1 плоскость симметрии. Двусимметричное Д. очень редкое (гребневики) - 2 плоскости симметрии.
При гаструляции происх. сл. перемещения кл., в рез. кот. часть его попадает внутрь ранее 1сл. зародыша (бластулы) и выстилает его ст, кот. становится 2сл. (Гаструла) + полость (гастроцель ), сообщ. с нар. средой при помощи отверстия - бластопора . Нар. ст. наз. эктодермой, внутр. - энто-. Это первичные зародышевые листки. В состав энто-, реже экто- вначале вх. и материал ср. листка - мезодермы. К концу Г мезодерма выдел. в самост. слой, и зародыш из 2сл в 3сл. 4 осн. мех-ма : Инвагинация (часть ст. 1сл. зародыша вворачивается внутрь и обр. энто-); эпиболия (относ. крупные, богатые Ж кл. обрастают мелкими и оказываются внутри, обр. энто-); иммиграция (отд. кл. внутрь зародыша и размещение их под поверхн. сл.); деламинация / раздел. кл. || пов-сти, due to 1сл. ст. в 2сл.
Нейруляция – обр. зачатка НС - Н пластинки и ее замыкание в Н тр. Во время Н в составе зарод. листков вычленяются зачатки отд. с-м органов. Зародыш - нейрулой.
Органогенез достигается в осн. за счёт разнообр. кл. перемещений и дифференц. самих кл. Произв. экто- выполняют покровную, чувств. и двиг. ф; из них - НС, кожный покров и обр. из него производные, эпителий пер. и зад. отд. пищевар. с-мы, соед/тк. основа кожи, пигментные кл. и висцеральный скелет. Энто- обр. выстилку кишечной полости и обеспеч. пит. зародыша; из неё возн. слизистая об. пищевар. с-мы, пищевар. ж-зы, органы дых. Мезо- осущ. св. м/у частями зародыша и вып. опорную и трофическую ф; из неё обр. органы выделения, половые органы, кров. с-ма, серозные об, выстил. целом и одевающие внутр. органы, мышцы; у позвоночных обр. + скелет.
76.
Система пищеварения. Регуляция пищеварения.
| Отдел | Функции |
| Ротовая полость | Прием пищи и ее пережевывание |
| Глотка | Проглатывание |
| Пищевод | Соединяет глотку с желудком |
| 12перстная кишка Печень (желчь) Поджел. железа (панкреатический сок) | Переваривание и всасывание Эмульгирование жиров Переваривание крахмала, белков и жиров |
| Подвздошная кишка | Завершение пищевар. и всасывание прод. |
| Толстый кишечник | Всасывание воды |
| Прямая кишка | Формирование каловых масс |
^ Р
 отовая полость
. На языке вкусовые луковицы. Слюна секретируется 3 парами слюнных и множеством щечных желез, расположенных в слизистой полости рта. Слюна – водянистый секрет, сед. амилазу и лизоцим, а также NaCl, бикарбонаты, фосфаты, карбонаты, ионы Ca, K, Mg и слизь. Слизь облегчает проглатывание пищи, смачивает и обволакивает. В конечном счете полужидкая, частично переваренная пища склеивается муцином и проглатывается. Пищевод
. 25 см. Им. мышечные ст. (вверху П-П+гладкая; внизу – тока гладкая), выстланные многосл. плоским эпителием, содержащим слизистые железы. Желудок
. Мышечная оболочка состоит из 3 слоев гладких мышц (нар. –продольный, промежуточный – циркулярный и внутр. – косой). Толстая слизистая об. Ж обильно снабжена эпителиальными кл, секрет. слизь, и покрыта многочисленными Ж ямками
. В этих ямках – главные и обкладочные кл., а также ферменты и соляная к-та. Слизь создает барьер м/у слизистой Ж и желудочным соком. 2 сфинктера: кардиальный
(верхний, кардиальный, отдел Ж, св. с пищеводом) и пилорический
(нижний, св. с 12п кишкой). Кислота (0,04-0,05%) оказывает бактерицидное дей-вие, разрыхляет ткани, размягчает волокна и способствует превращению пепсиногена
в пепсин
и прореннин
в ренин
. Пепсин расщепляет Б на полипептиды, а также превращает новые молекулы пепсиногена в пепсин (аутокатализ). Ренин в присутствии ионов Са вызывает коагуляцию растворимого Б молока – казеиногена
– и превращение его в нерастворимую кальциевую соль, кот. перевар. пепсином. Химус
– кашица. Внутр. фактор Ж
– необходим для всасывания молекул витамина В12.Тонкий кишечник
. Из 2 отделов: 12п.(20 см) и подвздошная (5 м) кишки. Стенка ворсинок обильно снабжена капиллярами и лимфатическими сосудами и сод. волокна гладких мышц. Они способны непрерывно сокращаться и вытягиваться (постоянный контакт с пищей). Слизистые кл. секретируют слизь. Расположенные в начале 12п кишки железы тоже секретируют слизь, а также щелочную жидкость (защита слизистой от поступ. из Ж кислоты). Слизистая тонкого секрет. ряд Ф, составляющих кишечный сок
(амилаза, мальтаза, лактаза, сахараза
и др.). Поджел. железа сост. из групп клеток – ацинусов, кот. секретир. ряд ферментов (амилаза, липаза, трипсиноген
и др.). Толстый кишечник
. Слизь + симбиот. бактерии
(а.к.
и витамины
, в том числе витамин К, кот. всасываются в кровяное русло). РЕГУЛЯЦИЯ
. Секреция пищевар. ферментов требует затраты Е, поэтому она происх. не постоянно. Секреция слюны
регулируется 2 рефлексами: безусловным (ч/з черепно-мозговые Н от пищи во рту) и условным (запах, вид, мысли). Секреция желудочного сока
протекает в 3 фазы: 1)нервная (пища во рту вызывает импульсы, кот. ч/з блуждающий Н передаются в Ж и стимулируют секрецию сока его слизистой – подготовка к А); 2)фаза растяжения (секреция сока стимул. растяжением стенок Ж); 3)гуморальная (пища в Ж стимулирует обр. в пилорическом отделе гормона гастрина
, кот. ч/з кровь дистигает слизистой др. отделов Ж и вызывает 4 часовую секрецию сока с высоким содержанием соляной к-ты. Энтерогастрон
секретируется на присутствие в пище ЖК → угнетает секрецию Ж, замедляет перистальтику и задерживает его опорожнение. Когда кислый химус поступает в 12п кишку и соприкасается с ее стенками, ее слизистая начинает секретировать кишечный сок
и 2 гормона: холецистокинин-панкреозимин
и секретин
.
отовая полость
. На языке вкусовые луковицы. Слюна секретируется 3 парами слюнных и множеством щечных желез, расположенных в слизистой полости рта. Слюна – водянистый секрет, сед. амилазу и лизоцим, а также NaCl, бикарбонаты, фосфаты, карбонаты, ионы Ca, K, Mg и слизь. Слизь облегчает проглатывание пищи, смачивает и обволакивает. В конечном счете полужидкая, частично переваренная пища склеивается муцином и проглатывается. Пищевод
. 25 см. Им. мышечные ст. (вверху П-П+гладкая; внизу – тока гладкая), выстланные многосл. плоским эпителием, содержащим слизистые железы. Желудок
. Мышечная оболочка состоит из 3 слоев гладких мышц (нар. –продольный, промежуточный – циркулярный и внутр. – косой). Толстая слизистая об. Ж обильно снабжена эпителиальными кл, секрет. слизь, и покрыта многочисленными Ж ямками
. В этих ямках – главные и обкладочные кл., а также ферменты и соляная к-та. Слизь создает барьер м/у слизистой Ж и желудочным соком. 2 сфинктера: кардиальный
(верхний, кардиальный, отдел Ж, св. с пищеводом) и пилорический
(нижний, св. с 12п кишкой). Кислота (0,04-0,05%) оказывает бактерицидное дей-вие, разрыхляет ткани, размягчает волокна и способствует превращению пепсиногена
в пепсин
и прореннин
в ренин
. Пепсин расщепляет Б на полипептиды, а также превращает новые молекулы пепсиногена в пепсин (аутокатализ). Ренин в присутствии ионов Са вызывает коагуляцию растворимого Б молока – казеиногена
– и превращение его в нерастворимую кальциевую соль, кот. перевар. пепсином. Химус
– кашица. Внутр. фактор Ж
– необходим для всасывания молекул витамина В12.Тонкий кишечник
. Из 2 отделов: 12п.(20 см) и подвздошная (5 м) кишки. Стенка ворсинок обильно снабжена капиллярами и лимфатическими сосудами и сод. волокна гладких мышц. Они способны непрерывно сокращаться и вытягиваться (постоянный контакт с пищей). Слизистые кл. секретируют слизь. Расположенные в начале 12п кишки железы тоже секретируют слизь, а также щелочную жидкость (защита слизистой от поступ. из Ж кислоты). Слизистая тонкого секрет. ряд Ф, составляющих кишечный сок
(амилаза, мальтаза, лактаза, сахараза
и др.). Поджел. железа сост. из групп клеток – ацинусов, кот. секретир. ряд ферментов (амилаза, липаза, трипсиноген
и др.). Толстый кишечник
. Слизь + симбиот. бактерии
(а.к.
и витамины
, в том числе витамин К, кот. всасываются в кровяное русло). РЕГУЛЯЦИЯ
. Секреция пищевар. ферментов требует затраты Е, поэтому она происх. не постоянно. Секреция слюны
регулируется 2 рефлексами: безусловным (ч/з черепно-мозговые Н от пищи во рту) и условным (запах, вид, мысли). Секреция желудочного сока
протекает в 3 фазы: 1)нервная (пища во рту вызывает импульсы, кот. ч/з блуждающий Н передаются в Ж и стимулируют секрецию сока его слизистой – подготовка к А); 2)фаза растяжения (секреция сока стимул. растяжением стенок Ж); 3)гуморальная (пища в Ж стимулирует обр. в пилорическом отделе гормона гастрина
, кот. ч/з кровь дистигает слизистой др. отделов Ж и вызывает 4 часовую секрецию сока с высоким содержанием соляной к-ты. Энтерогастрон
секретируется на присутствие в пище ЖК → угнетает секрецию Ж, замедляет перистальтику и задерживает его опорожнение. Когда кислый химус поступает в 12п кишку и соприкасается с ее стенками, ее слизистая начинает секретировать кишечный сок
и 2 гормона: холецистокинин-панкреозимин
и секретин
. 77. Система кровообращения и её регуляция.
С ердце окруж. околосерд. сумкой / перикардом
. Нар. слой кот. сост. из нерастяж. белой фиброзной тк., а внутр. – из 2 листков, висцерального
и париентального
. Висцеральный сращен с сердцем, а париентальный – с фиброзной тк. В щель м/у листками выделяется перикардиальная (серозная) жид-сть, кот. ↓ трение м/у ст. сердца и окруж. тканями. П и Ж соед. предсердно-желудочковым отверстием, кот. закрыто в левой половине двустворчатым
, а в правой – трехстворчатым
клапанами. Со ст. Ж к клапанам прикрепляются сухожильные нити
, начинающиеся от сосочковых мышц
Ж. Сосочковые мышцы вместе с сухожильными нитями удерживают клапаны и при сокращении сердца позволяют им открываться только в сторону Ж. Ст. сердца сост. из 3 слоев: внутреннего – эндокарда
, ср. – миокарда
и нар. – эпикарда
. Эпикард плотно срастается с миокардом. На ур. оснований аорты и легочного ствола эпикард переходит в перикард. Сердечшая мышца им. поперечную исчерченность. Кл. мышцы сердца при помощи особых перемычек соед. др. с др. Т.о., при раздражении возле места раздражения возникает волна сокращения, кот. быстро распространяется по всему сердцу. Серд. мышца не способна к длительному сокращению
: систола
(сокращение) и диастола
(расслабление). Стенки вен и артерий сост. из 3 оболочек (внутр. – эндотелий, ср. – мышечная, нар. – соед/тк), а капилляров из 1. Стенка вен тоньше артериальной: в ней слабо разв. мышечный слой и меньше эластических волокон.
ердце окруж. околосерд. сумкой / перикардом
. Нар. слой кот. сост. из нерастяж. белой фиброзной тк., а внутр. – из 2 листков, висцерального
и париентального
. Висцеральный сращен с сердцем, а париентальный – с фиброзной тк. В щель м/у листками выделяется перикардиальная (серозная) жид-сть, кот. ↓ трение м/у ст. сердца и окруж. тканями. П и Ж соед. предсердно-желудочковым отверстием, кот. закрыто в левой половине двустворчатым
, а в правой – трехстворчатым
клапанами. Со ст. Ж к клапанам прикрепляются сухожильные нити
, начинающиеся от сосочковых мышц
Ж. Сосочковые мышцы вместе с сухожильными нитями удерживают клапаны и при сокращении сердца позволяют им открываться только в сторону Ж. Ст. сердца сост. из 3 слоев: внутреннего – эндокарда
, ср. – миокарда
и нар. – эпикарда
. Эпикард плотно срастается с миокардом. На ур. оснований аорты и легочного ствола эпикард переходит в перикард. Сердечшая мышца им. поперечную исчерченность. Кл. мышцы сердца при помощи особых перемычек соед. др. с др. Т.о., при раздражении возле места раздражения возникает волна сокращения, кот. быстро распространяется по всему сердцу. Серд. мышца не способна к длительному сокращению
: систола
(сокращение) и диастола
(расслабление). Стенки вен и артерий сост. из 3 оболочек (внутр. – эндотелий, ср. – мышечная, нар. – соед/тк), а капилляров из 1. Стенка вен тоньше артериальной: в ней слабо разв. мышечный слой и меньше эластических волокон.
Сердечный цикл : 1) П в фазе диастолы, заполняются кровью; 2)Систола П; кровь выталкивается в Ж; клапаны открыты; сфинктеры полой и легочной вен открыты; 3) П расслабляются, Ж сокращаются; кровь выталкивается в аорту и легочную артерию; 4) клапаны аорты и легочной артерии закрыты; П начинают вновь заполняться кровью; Ж в фазе диастолы.
РЕГУЛЯЦИЯ : в продолговатом мозге имеется ряд участков, регулир. Дея-сть ССС, в том числе частоту сокращений сердца. От расположенного здесь кардиоингибиторного центра отходит пара блуждающих Н, сод. парасимпатические волокна. Поступающие по этим волокнам импульсы ↓ частоту серд. сокращений. В продолговатом м. также нах. уч. сосудо-двигательного центра , от кот. берут нач. Н симпатической с-мы. Импульсы, идущие по ним, ускоряют работу сердца. Совместная дея-сть волокон контролирует частоту серд. сокращений. К сердечно-сосудистым центрам продолговатого мозга подходят чувствительные Н волокна от Рs растяжения. Импульсы, поступающие от аорты замедляют работу сердца, в то время, как импульсы, поступающие от полых вен – ускоряют ее. При кол-ва крови в этих сосудах их стенки растягиваются, в результате чего число импульсов, посылаемых от них в СС центры продолговатого мозга.